

多巴胺 (成瘾元凶)
多巴胺是大脑中含量很丰富的儿茶酚胺类神经递质。多巴胺作为神经递质调控中枢神经系统的多种生理功能。多巴胺系统调节障碍涉及帕金森,精神分裂症,Tourette综合症,注意力缺陷多动综合症和垂体肿瘤的发生等。
多巴胺是一种神经传导物质,用来帮助细胞传送脉冲的化学物质。这种脑内分泌物和人的情欲、感觉有关,它传递兴奋及开心的信息。另外,多巴胺也与各种上瘾行为有关。阿尔维德·卡尔森(Arvid Carlsson)确定多巴胺为脑内信息传递者的角色,使他赢得了2000年诺贝尔医学奖。
简介
多巴胺(DA,或3-羟酪胺,3、4-二羟苯乙胺)是内源性含氮有机化合物,为酪氨酸(芳香族氨基酸)在代谢过程中经二羟苯丙氨酸所产生的中间产物。 又名儿茶酚乙胺或羟酪胺,是儿茶酚胺类的一种,分子式为C8H11NO2。是在中枢神经系统中存在特殊的多巴胺能系统,由黑质致密带发出的黑质纹状体束及黑质苍白球束,即属于多巴胺能神经通路,其神经介质是多巴胺,由神经细胞或突触本身合成。多巴胺除了作为去甲肾上腺素的前身外,还是维持锥体外系神经功能的重要神经介质。
多巴胺具有β受体激动作用,也有一定的α受体激动作用,能增强心肌收缩力,增加心排血量,加快心率作用较轻微,对周围血管有轻度收缩作用,升高动脉压,对内脏血管则使之扩张,增加血流量,使肾血流量及肾小球滤过率均增加,从而促使尿量及钠排泄量增多,能改善末梢循环,明显增加尿量,对心率则无显著影响,为其优于其他血管收缩剂或血管扩张剂之处,临床主要可用于各种类型休克。
锰中毒时,左旋多巴脱羧酶受到抑制,使左旋多巴脱羧为多巴胺及去甲肾上腺素的过程受阻,故在基底神经节中多巴胺及去甲肾上腺素的含量明显下降。
多巴胺虽早在1910年即已被合成,但与其密切相关的生物源儿茶酚胺(即肾上腺素和去甲肾上腺素)相比,由于相对较弱的拟交感神经活性而长期地被忽视了。一直到在动物组织内发现了α-多巴脱羧酶,多巴胺才作为正常人尿液中的组成成分而被观察。多巴胺在正常脑内的浓度至少与去甲肾上腺素一样高的事实启示,多巴胺除了作为去甲肾上腺素的前体以外,还可能有其他功能。 临床上常用的是盐酸盐和溴化氢盐。即使是多巴胺盐,在空气和光照下也容易被氧化。多巴胺溶液在弱酸性条件下比较稳定,而在碱性条件下则易被逐渐氧化。多巴胺的这一特性,近来已经被用于建立在体内和体外实验中都能敏感地测定多巴胺及其代谢产物的电化学技术。
含多巴胺的神经元,其细胞体主要分布在黑质、脚间核和丘脑下部等处。在这些区域多巴胺含量很高。多巴胺在机体内合成时以酪氨酸为原料。脑内的多巴胺主要是由黑质细胞来合成,这些多巴胺参与锥体外系统的活动,与躯体运动机能有密切关系。脑内多巴胺代谢失常时,可引起震颤性麻痹(帕金森震颤)。其症状是:全身肌肉紧张度增高,肌肉强直,随意运动减少,动作缓慢,面部表现呆板。患者静止时,上肢或下肢及头部震颤;情感激动时震颤增加;进行自主运动时震颤减少;入睡后震颤停止。中脑边缘部分的多巴胺神经元位于中脑脚间核,这些区域的多巴胺和精神活动的调节有关。有人认为精神分裂症是脑内这些部位的多巴胺过多的结果。由下丘脑弓状核发出的轴突到达靠近垂体的正中隆起,这些多巴胺神经元与垂体的某些分泌(特别是促腺性素的分泌)有关。
多巴胺受体
多巴胺受体
多巴胺受体是结合在膜上的供神经递质多巴胺识别的位点。多巴胺受体既存于中枢神经系统(CNS),也存在于外周。依据生化和药理学标准已将此受体分为二型。微摩浓度的多巴胺作用于D1多巴胺受体可刺激腺甘酸环化酶的活性。酚噻嗪类多巴胺拮抗剂如氟非那嗪作用非常强大,抑制多巴按D1受体效应只需纳摩水平,而丁酰苯类拮抗如像氟哌啶醇的药效则弱得多,在微摩波度才显示效应。具有多巴胺效应的麦角类则是D1受体弱的部分激动剂。相反,纳摩浓度的多巴胺激动剂作用于D2受体就能抑制由其他激素或者神经递质激活的腺苷酸环化酶活性。对D2受体来说,酚噻嗪类和丁酰苯类作用效应是纳摩级的,而且多巴胺效应的麦角则是作用强大的完全激动剂。苯酰胺类抗精神病药物,如舒必利作为拮抗剂其效力为纳摩到微摩级,但它们对D1多巴胺受体无作用。
D1和D2多巴胺受体都典型地发现于对多巴胺神经末梢是突触后的成分,两类受体也见于皮层-纹状体的末梢,在此多巴胺末梢与谷氨酸能末梢形成轴突-轴突型突触以调节谷氨酸的释放。重要的是,在多巴胺细胞胞体,树突和末梢都有多巴胺受体发现。这些自身受体既调节多巴胺的合成、释放,也调节神经元的冲动发放频率。从药理学角度看,这些受体似与D2受体性质相似。刺激脑内多巴胺受体产生的确切行为变化尚不清楚。多巴胺通过在锥体外运动系统中的作用肯定参与运动调节。当黑质纹状体多巴胺通路受损时,将导致帕金森病的运动功能丧失或运动不能的产生。多巴胺似乎也参与摄食和摄水的增强和调节。
D2多巴胺受体也存在于垂体前叶和中叶。在垂体前叶,多巴胺抑制催乳素细胞释放催乳激素。下丘脑弓状核中的多巴胺细胞体释放多巴胺入门脉系统,随血流入垂体前叶。在垂体中叶,多巴胺通过抑制由β2-肾上腺素能受体激活的腺苷酸环化酶活性以调节α-促黑激素(α-MSH)的释放。多巴胺受体也存在于外周:在交感神经末梢上的D2受体,可抑制去甲肾上腺素的释放;D1受体则主要存在于肾血管网上,受刺激时可引起血管扩张。两型受体还见于视网膜无长突细胞层,但其功能还不清楚。
受体的分布
在缺乏每种多巴胺受体亚型的特异配体之前,广泛应用原位杂交的方法来研究多巴胺受体mRNAs在脑内的分布。D1和D2受体基因在脑内表达广泛。D1-R主要表达于尾壳核,伏隔核,视束,脑皮层和杏仁核。除此之外,D1受体还在下丘脑被探测到。尽管在黑质致密部发现有D1配体与其结合,但没有探测mRNA存在。这一结果提示:D1-R在纹状体合成后通过纹状体黑质束转运到黑质。D5-R和D1-R相比表达局限,仅在海马,外侧乳头体核和下丘脑束旁核表达。D2-R mRNA主要在脑的CPu, OT和Acb中表达。在黑质和腹侧被盖部(VTA)也有表达,这些区域发出多巴胺纤维,提示着D2-R有突触前定位。相反,D1样受体只有广泛的突触后定位。分析D2样受体的两种亚型的mRNA表明D2L表达极为丰富。在脑外,D2-R mRNA还定位于视网膜,肾脏,血管系统和垂体。大鼠脑D3-R的mRNA分布仅限于隔核,下丘脑和丘脑,以及小脑中的某些区域。D3-R还分布于黑质致密部,提示它也存在突触前定位。在前额叶皮层,杏仁核,嗅球,海马,丘脑和中脑可见D4-R的mRNA高度表达。
应用多巴胺受体亚型特异抗体可对其在不同脑区进行细胞和亚细胞定位。D1和D5受体共同表达于前额叶皮层,运动前区,扣带和内嗅皮层,海马和齿状回的锥体细胞。电子显微镜证实D1和D5受体存在于前额叶皮层,海马的突触前和突触后,以突触后分布更常见。超微结构分析发现:D1和D5受体在人锥体细胞分布不同,D1受体集中在树突棘,D5受体集中位于树突轴。在嗅球,D1受体限于内颗粒层和内从层;在杏仁核,其限于中介核和基底外侧核。在尾状核,D1和D5受体大多数位于中等大小的GABA能神经元。D5受体也存在于大的胆碱能中间神经元。超微结构分析发现: D1受体存在于非对称性突触的突触后树突棘部, D1和D5受体位于多巴胺终端特征性小突触的突触后树突上,以及突触前D1和D5受体位于形成非对称性突触的轴突上。已明确D1受体位于脚内核和黑质网状部,这些区域并未检测到D5受体。这一结果提示:如果D1和D5受体在尾状核中等大小多棘神经元共存,只有D1受体被转移到纹状体靶区终端。尽管D1和D5受体有着相似的药理学特性,但细胞和亚细胞定位不同提示它们在功能上可能不同。用特异抗体的免疫组化方法显示:D2受体存在于纹状体中等多棘神经元,在棘突和棘头的分布比胞体密集。与D1受体的共存非常罕见。D2免疫反应终端形成的对称性突触多于非对称性突触。D2受体存在于黑质致密部的核周体和树突,和其他纹状体投射相比,更集中于苍白球外段。D2受体免疫阳性反应还可在嗅球的颗粒层和内从层以及杏仁核见到。免疫组化和电子显微镜揭示了D4受体存在于大脑皮层和海马的锥体和非锥体神经元,它们已被证实是GABA能中间神经元。在大脑皮层和海马,D4受体调控GABA的传递。D4受体也在苍白球片段,黑质网状部以及丘脑网状核的GABA能神经元内被发现。
多巴胺的功能
运动
多巴胺对运动控制起重要作用,帕金森病是由于多巴胺能神经元变性引起严重的多巴胺减少所致。多巴胺拮抗剂和激动剂应用的研究表明了多巴胺受体在运动控制中的重要作用如:大鼠的前进,后退,僵直,吸气和理毛功能。通常激动剂提高多巴胺的运动功能,拮抗剂作用相反。已明确了在决定向前运动中的D1和D2受体有相互促进作用。
同时刺激D1受体可使D2受体激动剂产生很大的运动刺激。对多巴胺受体(D1-4)基因突变鼠的行为分析可提供每一种运动亚型。与药理学研究结果相反,D1-R突变鼠的运动功能和野生型鼠相比未受影响,说明了在调控自主运动过程中不同多巴胺受体之间相互作用的复杂性。D2-R敲除鼠出现明显的运动功能受损,如运动减少,不协调或后退不能。D3-R突变鼠出现运动过度的运动亚型,与应用药理学方法给予D3-R激动剂或拮抗剂的结果一致。D4-R突变鼠的运动功能也受到影响。
药物成瘾
中脑边缘多巴胺系统与药物成瘾包括阿片,可卡因,安非他命和乙醇等造成的精神运动效应以及奖励机制的控制有关。可卡因和安非他命通过阻断多巴胺转运子(DAT)的活动,翻转多巴胺的运输增加多巴胺在突触间隙的释放。然而,阿片在纹状体,前额叶皮层,视束,伏隔核,下丘脑内侧基核和杏仁核对多巴胺的释放起抑制效应。已证明破坏伏隔核,或用D1或D2-R拮抗剂阻断多巴胺受体,可削弱吗啡,可卡因和安非他命引起的运动过度和奖励效应。D1-R敲除鼠行可卡因治疗后,不出现经典的运动过度和刻板的行为效应。在地点选择性实验模型中,D2-R敲除鼠和野生型鼠相比缺乏对吗啡的条件反射。D3-R突变鼠对可卡因的敏感性增加。D4-R突变鼠对乙醇,可卡因,安非他命引起的运动活动刺激更加敏感。
学习记忆
普遍认为中脑边缘多巴胺系统和中脑皮层多巴胺系统在学习记忆中发挥作用。有报道,猴A10区多巴胺能神经元涉及学习认知行为中基本注意力和动机过程中的冲动活动的短暂变化。
D1和D2受体可调节多巴胺在学习记忆中的作用。许多实验表明多巴胺通过D1-R调控前额叶皮层(PFC)神经元的活动和工作记忆过程。D2和D3-R基因缺陷鼠和野生鼠相比,出现延迟交替作业有关空间记忆缺陷,预先给予METH(metaphetamine)处理的突变鼠的空间学习能力未受损,说明D2-R调节学习记忆机制是通过D1-R介导的。研究发现,健康志愿者口服D2-R激动剂溴隐亭,可提高空间学习记忆的能力;而口服D2-R拮抗剂,可出现空间识别能力损害,且呈剂量依赖性下降。
脑老化
随着年龄的增长,认知功能将随之下降这一事实,在人类和灵长类中都得到了很好的证明。近年来有研究提供了直接的证据,证明了多巴胺功能下降,认知功能下降和脑老化的明显相关性,对同年龄的志愿者进行对比性研究,其结果表明,多巴胺水平与认知功能呈正相关,与年龄呈负相关。
长时程增强(LTP)
长时程增强为目前神经科学家所认同的学习记忆的细胞模式。在自由活动的大鼠海马齿状回颗粒细胞层植入慢性记录电极,在中前通路植入刺激电极,同侧脑室植入套管给药,发现预先给予D2样受体阻滞剂remoxipride可抑制海马弱的LTP(<4 h),说明D2样受体通过调控体内海马LTP和去极化涉及学习记忆的相关过程。对培养的皮层纹状体细胞脑片进行细胞内记录研究,发现用内源性多巴胺或外源性多巴胺激动剂激活多巴胺D1/D5受体对皮层纹状体通路的LTP的诱导是必需的。
多巴胺代谢途径
合成和储存
络氨酸由饮食蛋白提供,或由苯丙氨酸经肝脏苯丙氨酸羟化酶转换而成,经氨基酸转运体入脑,多巴胺神经元,经胞浆酪氨酸羟化酶转换成二氢苯丙氨酸(左旋多巴),再由芳香氨基酸脱羧化酶(多巴脱羧化酶)转换成多巴胺。胞浆多巴胺转运至囊泡,囊泡多巴胺浓度为0.1M。
释放
当动作电位到达时,膜蛋白构造改变,允许Ca2+流入,囊泡与神经末梢或树突融合,通过胞吐作用将多巴胺释入突触间隙。有两种释放方式:一种是间断性释放,即动作电位到达时一过性释放多巴胺,然后快速回收入神经元;一种是持续性释放,即低水平持续释放多巴胺,此时的多巴胺水平不足以激动突触后膜多巴胺受体,只能激动突触前膜多巴胺自身受体,抑制间断性释放。
回收和代谢
神经末梢经转运体或膜内外浓度差,将多巴胺回收入神经末梢,以供再利用。神经胶质细胞和非多巴胺神经元一定程度上也回收和代谢多巴胺,代谢酶包括单胺氧化酶、儿茶酚胺甲基转移化酶和醛脱氢酶。
中科院上海生命科学院神经科学所发现一种小G蛋白的调节因子Vav2能够通过调节多巴胺转运体在质膜的分布,从而显著改变多巴胺“回收泵”系统的转运效率。如果将Vav2基因敲除,“回收泵”功能异常提升,就会使大脑伏隔核多巴胺的含量明显升高。为了寻找控制多巴胺“回收泵”的“开关”,研究人员利用分子生物学实验手段筛选到胶质细胞源性神经营养因子GDNF的受体Ret。研究结果显示,GDNF和Ret可作为拨动和调节多巴胺“回收泵”的“开关”而起作用。当这套“开关”失灵(如Ret基因敲除)的时候,动物呈现类似于Vav2基因敲除小鼠的表现。过去一般认为,GDNF及其受体Ret主要是掌管多巴胺能神经元的存活,因此,他们的这一发现拓展了人们对神经营养因子 GDNF作用的传统认识。
多巴胺受体
能结合SCH23390的称为多巴胺D1受体, D1受体与兴奋性核苷酸结合蛋白复合物相互作用,激活腺苷酸环化酶系统;能高亲和性结合丁酰苯类药物(螺哌隆和氟哌啶醇)的称为D2受体,D2受体与抑制性核苷酸结合蛋白复合物相互作用,抑制腺苷酸环化酶系统。重组DNA技术将这两种受体再分,多巴胺D1样受体再分为D1和D5受体,多巴胺D2样受体再分为D2、D3和D4受体。
通路
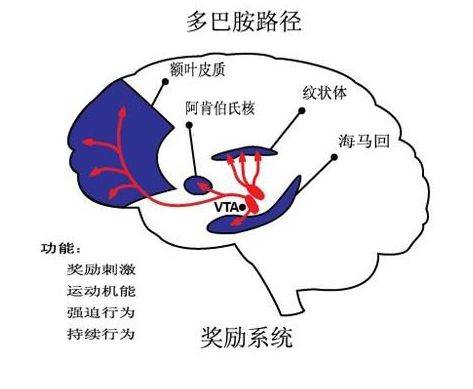
多巴胺神经元位于中脑与间脑交接处,有3条通路。第一条是黑质-纹状体通路,第二条是中脑-边缘通路,第三条是中脑-皮质通路。
(1)黑质-纹状体通路
黑质-纹状体通路起自黑质致密带,投射至尾核-壳核,该通路激活能增加运动,动物黑质损害能引起运动迟缓和反应时间延长。抗精神病药阻断多巴胺D2受体,表现为锥体外系反应。
(2)中脑-边缘通路
该通路起自腹侧被盖区外侧神经元,多数投射到伏膈核和嗅结节,少数投射到中膈、杏仁核和海马,激动时引起愉快体验,抑制则引起心绪不良。当睾酮升高时,增加 γ-氨基丁酸(GABA)回收,多巴胺突触前膜上的GABAA受体激动不足,导致多巴胺脱抑制性释放,男、女性唤醒和性享受增加。
(3)中脑-皮质通路
该通路起自腹侧被盖区内侧神经元,投射至前额内侧、鼻周皮质。当机体暴露有意义的新刺激时,中脑腹侧被盖区释放多巴胺,引起警觉,当反复暴露该刺激时,机体熟悉后,就不再释放多巴胺。激动额叶皮质与纹状体复合物环路时,反映了兴趣或厌恶的强度,即精神动力。多巴胺增强这种精神动力,多巴胺阻断剂则削弱这种精神动力,表现为情感淡漠和意志缺乏,称为继发性阴性症状。
额叶皮质多巴胺升高时,激动D1受体能改善注意、目标指向行为、犒赏和学习。苯丙胺阻断多巴胺回收,从而激动D1受体,改善认知功能;典型抗精神病药阻断D1受体,损害认知功能;不典型抗精神病药阻断多巴胺神经纤维突触前膜上的5-HT2A受体,引起多巴胺脱抑制性释放,改善认知功能。
(4)中脑-边缘-皮质多巴胺通路
尽管描述中脑-边缘通路的多巴胺神经元在腹侧被盖区外侧,中脑-皮质通路的多巴胺神经元在腹侧被盖区内侧,但两者还是有相当大的重叠,故有时统称中脑-边缘-皮质多巴胺通路,其激活能增加精神动力。随着年龄的增加,多巴胺受体减少,这可能解释老人的动力和快感不如青少年强。
(5)反馈通路:纹状体-黑质神经元和中脑局部环路
神经元释放GABA,激动GABAA和GABAB受体,抑制多巴胺神经元。苯二氮 类药物激活GABAA受体,通过抑制多巴胺神经元而恶化抗精神病药的锥体外系反应,另一方面,通过肌肉松弛效应而改善锥体外系反应,如此一反一正,故苯二氮类药物未推荐为治疗锥体外系反应。
快感引起机制
基底前脑
伏膈核富含多巴胺神经元;分为两部分:中央为“核”,边缘为“壳”,壳是杏仁核的延伸部分。活体微透析分析发现,天然犒赏(如食物、水、性和其他愉快性刺激)增加伏膈核多巴胺释放。相反,杏仁核促进害怕行为,而前额皮质则调节总动机,决定行为反应的强度。弗洛伊德爱的本能、恨的本能和自我,分别可用这三个解剖部位解释,这三个解剖部位的关系可用一个人低头作揖来模拟,双手抱拳相当于伏膈核核部,两前臂相当于伏膈核壳部,两上臂相当于杏仁核,头部相当于前额皮质,可以这样记:爱的中枢(伏膈核)在中间受到呵护,恨的中枢(杏仁核)在双侧护卫,上端有前额皮质调控。这三个解剖部位加上嗅结节,称为基底前脑。
犒赏通路
多巴胺犒赏通路是指腹侧被盖区投射至基底前脑的内侧前脑束。笼统地讲,是从中脑被盖区投射到前额皮质,激动释放多巴胺,经前额皮质-伏膈核核部谷氨酸通路激活伏膈核,引起寻药行为。当抑制中脑被盖区、前额皮质和伏膈核这三个环节的任何一个环节时,都能阻止觅药行为。
犒赏递质
有两种,一种是多巴胺,与鼓励和准备获得犒赏相关联,如动物交媾前的交配仪式、人类调情的愉快、用药前线索(如看电影上的人注射海洛因)引起的愉快渴望,表现为一阵激动感、迫切感或渴望感;另一种是阿片类物质,与圆满犒赏相关联,包括镇静、休息和“极乐感”。如急性饮酒能刺激 β-内啡肽释放,引起圆满犒赏。
多巴胺激联
成瘾性毒品刺激多巴胺过量释放→激动多巴胺D1受体→兴奋性核苷酸结合蛋白复合物增多→腺苷酸环化酶增多→环-磷酸腺苷增多→蛋白激酶增多→环-磷酸腺苷反应成分结合蛋白磷酸化→诱导立即早期基因产物(如cFos)→短期神经形成改变→急性犒赏效应。
不快引起机制
当犒赏通路多巴胺能低下时,引起快感缺失(不能体验到愉快)、情感淡漠(缺乏动机)和心绪不良(感到不满意、不幸福、痛苦、激惹和紧张不安)。
快感缺失
(1)心境恶劣
假定素质性伏膈核多巴胺能低下导致持续的快感缺失,表现为持续2年以上的心境恶劣。由于青少年时期的基础多巴胺能极低,故青少年起病。当给予犒赏刺激时,多巴胺一阵释放,引起犒赏效应,心境恶劣暂时缓解;犒赏刺激消失,多巴胺再度低下,再次陷入心绪恶劣。即使犒赏刺激持续存在,由于不再新颖,多巴胺不再释放增加,病人也会再次心绪恶劣,这可解释心境恶劣缘何波动,缘何有不足2个月的缓解期。
心境恶劣有一个特点,就是无论用什么抗抑郁药,起初多少有效,过一段时间后效力减退,可能是这些药物作为一种新颖刺激(安慰剂样效应),引起多巴胺一阵释放,时间一长,该刺激不再新颖,多巴胺释放不再释放增加,再次陷入心境恶劣状态。
(2)重性抑郁症
当犒赏通路多巴胺能非常低下时,引起严重的快感缺失,这是抑郁症的一项核心症状,重性抑郁症通过物质使用障碍(Substance useddisorder)而增加伏膈核多巴胺能,改善快感缺失,其物质使用障碍率比普通社区人群为高(27%:17%)。丁氨苯丙酮抑制去甲肾上腺素和多巴胺回收,治疗三环抗抑郁药难治的抑郁症有效。
(3)精神分裂症
一些精神分裂症病人服用抗精神病药、活动缺乏、失业、生活在低社交环境中,引起多巴胺能不足,表现为快感缺失或心绪不良,病人选择物质滥用,增加多巴胺能,缓解快感缺失和心绪不良。故精神分裂症的物质滥用率是常人的4倍,包括酒中毒和大街药物滥用,如果服抗精神病药期间主观幸福感高,则滥用物质率低。
情感淡漠
(1)淡漠形成
如果遗传或围产期因素导致儿童突触密度低,到青少年期再修剪一次,导致突触联系过度减少;或原来突触密度不低,但青少年期过度修剪,导致突触联系过度减少。当额叶皮质多巴胺突触联系过度减少时,表现认知障碍(注意和语言表达缺损)、阴性症状(包括情感淡漠)和快感缺失/心绪不良。额叶皮质多巴胺突触联系过度减少还导致皮质-边缘谷氨酸-γ-氨基丁酸通路功能低下,该通路本来能抑制中脑-边缘多巴胺能通路,当功能低下时,中脑-边缘多巴胺能通路功能亢进,多巴胺释放增加,过度激动多巴胺D2受体,引起阳性症状(如幻觉妄想)。
(2)预报因素
同源相关假说认为,一种原因(多巴胺能不足)依赖于程度引起多种结果(快感缺失/心绪不良、情感淡漠和阳性症状),则多种结果之间应有相关性。故青少年期的快感缺失可能预示其易情感淡漠和阳性症状。
心绪不良
(1)主观体验
精神药物可引起三种主观体验:正性、中性和负性,可卡因、苯二氮 类药物和许多非法药物引起过强的正性体验(“刺激”或“高潮”),促进其反复使用,引起依赖和滥用;抗抑郁药引起中性或正性体验,促进治疗依从性;抗精神病药引起负性体验(如心绪不良),导致治疗不依从。
(2)生化机制
精神分裂症伏膈核多巴胺基础水平就低,抗精神病药阻断伏膈核D2受体,引起心绪不良,发生率10%~60%。初步证据表明,当占领D2受体不足60%时,病人仍处精神病状态,可伴负性3主观幸福感;当占领60%~70%时,主观体验很理想,可增加治疗依从性和生活质量;当高于70%时,则引起心绪不良。
(3)症状影响
有学者提出,静坐不能和运动不能决定了抗精神病药引起心绪不良的程度,即使是轻微的静坐不能,也能引起心绪不良。一位运动不能的女病人说,我服药后像一个老太,这对我的形象有重要影响。同样,过度镇静也可引起心绪不良。
(4)早期预测
有研究表明,住院精神分裂症病人服氯丙嗪试验剂量,如出现心绪不良,预示后来疗效差。对氨砜噻吨的研究表明,如首服剂量4~24小时内出现心绪不良,则病人可能立即不肯服药,进而预后差。因此,当给病人选择抗精神病药时,应考虑病人既往的主观体验报告。
(5)导致后果
心绪不良影响生活质量和功能状态,恶化治疗关系,导致服药不依从,后者导致疾病复燃率升高,预后差,甚至引发自杀。不典型抗精神病药的心绪不良少得多。机制可能是:不典型抗精神病药阻断5-HT2A受体,引起多巴胺脱抑制性释放,部分抵消D2受体的阻断效应。一项为期2年的研究表明,服利培酮组比服氟哌啶醇组的主观体验好,伴发的焦虑和抑郁少。尽管奥氮平引起的静坐不能较少,但比氟哌啶醇组的主观体验无明显优势。
多巴胺临床药物
作用与用途
可激动交感神经系统肾上腺素受体和位于肾、肠系膜、冠状动脉、脑动脉的多巴胺系统。效应与剂量有关:小量时,每分钟按体重0.5~2μg/kg使肾及肠系膜血管扩张;而小到中量,每分钟按体重2~10μg/kg对心肌产生正性应力作用;大量时,每分钟按体重大于10μg/kg,使肾血流量减少,收缩压及舒张压增高。适用于洋地黄及利尿药无效的心功能不全。用于心脏手术、肾功能衰竭、充血性心力衰竭、心肌梗死、内毒素败血症、创伤等引起的休克综合征及补充血容量效果不佳的休克。
用法与用量
成人常用量:静滴开始时按每分钟1~5μg/kg,10分钟内以每分钟1~4μg/kg的速度递增而达极佳疗效。慢性顽固性心衰可每分钟按0.5~3μg/kg可生效。闭塞性血管病变患者开始按每分钟1μg/kg速度静滴,渐增直到每分钟20μg/kg,以达疗效。如危重病例可先以每分钟5μg/kg滴注,然后增至每分钟20~50μg/kg。
注意事项
应用本品前应先纠正血容量,同时监测血压、心率、心律、尿量。外周血管灌流情况,休克纠正时应即减慢滴速,不可突然停药。防止药液外漏,选用粗大静脉。常见不良反应有胸痛、呼吸困难、心律失常(快而有力)、乏力、头痛、恶心、呕吐,长期使用可能导致局部坏死或坏疽,过量时发生严重高血压。对本品过敏者、孕妇、小儿慎用;闭塞性血管病、肢端循环不良、频发室性心律失常者慎用。嗜铬细胞瘤患者不宜使用。α受体阻滞剂、β受体阻滞剂可拮抗本品的作用;与硝酸酯类合用,互相减弱作用;与硝普钠、异丙肾上腺素、多巴酚丁胺合用时注意心排血量的改变;与全麻药合用时引起室性心律失常。本药可加强利尿剂的作用;与苯妥英钠联用时需交替使用;而与三环类抗抑郁药或单胺氧化酶合用可增强和延长本品的效应。
多巴胺的临床应用
休克
微循环动脉血灌流急剧减少,致重要生命器官因缺氧而发生功能和代谢障碍,是各种休克发生发展的共同规律。因此,休克的治疗应着重于尽快改善微循环,而不应单纯追求一个“满意”的血压。休克的恢复取决于微循环的改善,而不单纯取决于提升血压。在治疗休克时除常规扩容、积极处理原发疾病、纠正酸碱平衡失调外,根据病情需要及时应用DA,对于休克早期应该给予中小剂量的,使肾、脑、肠等重要器官的血管扩张,使有限的血流重新分配,同时又能使心输出量有所增加,血压适当提高。尤其适用于伴有心肌收缩力减弱、尿量减少而血容量已补足的休克患者。而对于顽固性低血压或感染性休克时,在使用大剂量效果仍不佳时,应及时改用血管收缩剂去甲肾上腺素。
心力衰竭
心力衰竭的主要特点是体液储留,导致肺淤血和外周水肿,造成呼吸困难和乏力,限制了运动耐量。因此有效改善体液储留可缓解患者的心力衰竭症状。首先对该类患者应用利尿剂治疗是必不可少的组成部分,而且必须尽早应用。另外在使用扩血管药物如硝普钠治疗心力衰竭时,联用DA可以起到协同、互补作用。因为在加强心肌收缩力而使心排出量有更大程度增加的同时,又可纠正在应用硝普钠时因前负荷减少而心排出量降低的情况,加之可维持血压,使硝普钠的使用范围扩大,不必担心血压降低不良反应。
肾脏“保护”
传统观念认为小剂量即所谓“肾剂量”DA有利尿保护肾脏作用,其机制为:(1)兴奋近曲小管血管上的受体产生肾血管扩张作用,增加肾血流量及肾小球滤过率,促进排钠、利尿;(2)抑制近端肾小管、髓拌升支以及皮质集合管处的钠-钾-ATP酶活性产生利尿作用;(3)使具有拮抗抗利尿激素作用的前列腺素E2产生增多而利尿;(4)激动β1受体,增加心输出量,增加肾灌注而增加尿量。
支气管哮喘
支气管哮喘是一种气道慢性炎性反应,临床上表现为反复发作性喘息、呼气性呼吸困难、胸闷或咳嗽等症状,常在夜间和或清晨发作、加剧,常常出现广泛多变的可逆性气流受限,多数患者可自行缓解或经治疗缓解。DA可直接兴奋气道平滑肌和肥大细胞膜表面的受体,舒张支气管平滑肌,减少肥大细胞和嗜碱性粒细胞脱颗粒和介质的释放,降低微血管的通透性。另外小剂量DA对外周血管有选择作用,可使肾血管、肠系膜血管、冠状动脉、脑血管扩张,使微循环得到改善,心脏前后负荷减轻,起到强心、利尿,保证重要器官血流灌注的作用。
滥用影响
D2受体功能低下
正电子发射计算机断层扫描研究显示,非药物滥用者服利他林(阻断多巴胺转运体),引起快感者脑D2受体基础水平低,不引起快感者脑D2受体基础水平高。提示当D2受体基础水平低时,提高多巴胺易引起快感。在现实中,基础多巴胺D2功能低下导致快感缺失,他们会通过暴食、性欲过强、赌博、冒险行为和服成瘾药物(如可卡因、苯丙胺、酒精、大麻、阿片类物质和尼古丁等)刺激多巴胺系统,以保证快感。
增加多巴胺能
阿片和大麻激动μ受体,从而增加多巴胺能;大麻的活性代谢物是 Δ-9-四氢大麻,通过刺激腹侧被盖区而增加伏膈核和前额皮质的多巴胺释放;酒精和苯环乙哌啶通过拮抗N-甲基-D-天门冬氨酸受体而使多巴胺脱抑制性释放;尼古丁通过激动尼古丁受体而增加多巴胺释放,苯丙胺刺激多巴胺释放;可卡因阻断多巴胺回收;均增加多巴胺能,引起快感。
易感药物滥用者
在青少年时期,中脑-边缘通路的多巴胺基础水平低下,这种低下可引起厌烦和不满意。当药物诱发多巴胺释放时,其释放量比成人为多,造成的犒赏效应比成人为大,故易感药物滥用。习惯性服用滥用药物后,中脑-边缘多巴胺通路发生耐受,同等药量不再引起多巴胺释放增加,快感不再出现,需要更大药量才能引起多巴胺释放增加,再度出现快感,此称药效耐受。当滥用药物撤退时,多巴胺水平低于基线水平以下,引起快感缺失和心绪不良,强制病人继续用药,引起药物依赖。
增加精神症状
许多研究显示,大麻使用障碍的精神分裂症病人治疗合作性低,精神病阳性症状更多,再住院率高,但低剂量大麻能缓解焦虑和抑郁。这从机理上可得到解释,因为大麻增加多巴胺能,而多巴胺D2受体过度激活能恶化精神分裂症阳性症状;低剂量大麻轻度增加多巴胺能,引起快感,故缓解焦虑和抑郁。多巴胺激动“犒赏”通路,故拟多巴胺药物可治疗某些药物(如可卡因)依赖。
聚多巴胺(PDA)
制备方法
多巴胺作为反应单体,碱性条件下(pH>7.5),氧气为氧化剂,反应过程不需要复杂的实验仪器和步骤,当多巴胺(通常选用多巴胺盐酸盐)在避光有氧的碱性条件下,即可发生聚合,在聚合的过程中溶液的颜色会随着时间的推移逐渐发生变化,颜色由无色变为棕色后变为黑色。在碱性条件下,多巴胺单体邻苯二酚去质子氧化后可以形成多巴胺醌,但多巴胺醌的结构不稳定会被继续氧化,发生分子内重排,形成深褐色的聚多巴胺。如果将聚多巴胺涂覆在不同的底物表面,只需将底物放在反应容器中轻轻振荡。一定的反应时间后即可在底物表面生成一层均匀致密的聚多巴胺涂层。多巴胺单体浓度控制在2g/L以上,形成的聚多巴胺薄膜厚度可通过反应时间的长短和多巴胺单体的浓度决定。研究表明,聚多巴胺薄膜的很大厚度为50nm,多巴胺单体的浓度和反应时间超过某一阈值后就不会再对膜的厚度造成影响。目前,除了溶液氧化法,还有酶氧化过程,利用酶催化聚合物合成。
聚多巴胺的应用
(1)聚多巴胺的结构特性
在不同的反应条件下,聚多巴胺的微观结构也是不一样的,虽然都属于黑色素材料,但是通过电镜微观观察,聚多巴胺(PDA)可以呈纳米带状、纳米球状等不同形貌,不同结构的聚多巴胺材料随着比表面积不同对污染物的吸附能力不尽相同,这要根据实际污水处理标准进行合理材料选择制备。聚多巴胺稳定性好,水分散性强,生物相容性优秀。常温下即可生成。制备不同结构特征的聚多巴胺微/纳米球可以通过控制氧化自聚的反应条件如多巴胺初始浓度、缓冲溶液的类型、反应时间等。聚多巴胺微/纳米球中含有大量的氨基和酚羟基等基团,由于氨基和酚羟基可以与金属离子结合,所以聚多巴胺微/纳米球也会被应用在处理水污染中的重金属离子。此外,聚多巴胺微/纳米球可作为金属离子的还原剂,制备金属纳米材料。
(2)聚多巴胺对膜的改性
研制一种表面能高、具有亲水性的分离膜用来处理废水中的污染物是科学家在膜技术领域致力的方向之一。实验表明在碱性有氧避光的条件下多巴胺能够在各种基底表面发生氧化聚合反应,形成一层致密且具有强黏附作用的复合层薄膜,在涂覆有聚多巴胺的薄膜表面接枝各种有利于目标污染物特异性吸附的改性分子或聚合物可以实现复合膜的功能化。
这种薄层复合纳滤膜在海水淡化方面具有很好的应用前景。用膜吸附水环境中的污染物也属于吸附法,制备各种复合膜来处理污水也是环境学家致力于研究的方向之一,用聚多巴胺不仅可以直接作为枝接物,还可以用作生物胶来连接不容易和膜黏附的化学物质。
(3)聚多巴胺对纳米材料表面修饰
聚多巴胺除了可以修饰各种固体基底材料和膜材料之外,还可以对纳米材料进行修饰。对于纳米材料,多巴胺可以进行普通的涂覆作用,也可以进行二次功能化修饰。纳米材料作为环境领域的一类新型吸附剂,具有很多优点,将聚多巴胺涂覆在纳米材料表面可以实现这类吸附材料在环境治理的更大应用能力。
聚多巴胺(PDA)可通过多巴胺(DA)的自氧化聚合涂覆在多种基体表面。由于多巴胺具有黏附性、还原性以及生物相容性,多巴胺及其衍生的复合材料逐渐被应用在了各个领域,其中在环境领域,可以将多巴胺应用在膜技术和污染物的吸附等方面。
传统的膜技术和吸附对需要改性的基底有较高的要求,而且在修饰过程中有可能对环境造成二次污染,但是,多巴胺的材料功能化会避免这种情况,整个制备过程也会是一个绿色环保、简单易行、功能稳定的途径。通过对固体、膜或者纳米材料等基底的表面改性制备出新型多功能的材料应用在水处理当中,大大提高了吸附剂的吸附性能。如今已经有很多成功的例子可以证明这种材料在环境吸附中的重要地位,而且人们还在不断探索它的创新应用。
国内对多巴胺在水环境处理的应用方面已经做出了许多科研成果,但是其只是停留在基础研究阶段,将多巴胺应用在实际的污水处理并将其产业化生产还不是非常广泛。多巴胺在环境领域可以给人类带来意想不到的作用。在国内外,多巴胺虽然已经广泛地应用在各种材料功能化上面,但其聚合机理并不能准确把握,依然停留在探索阶段,需要对其反应动力学及中间体的结构进行进一步研究,以拓展其应用领域。
-
01/26
2021
儿童焦虑症产生的原因是怎样的?家长该怎样做?
儿童焦虑症(anxiety disorder of childhood)是最常见的情绪障碍,是一组以恐惧不安为主的情绪体验。可通过躯体症状表现出来,如无指向性的恐惧、胆怯、心悸、口干、头痛、腹痛等。婴幼儿...

-
01/21
2021
K粉对于男性和女性的伤害一样嘛?
K粉对男性的危害:对男性而言,长期吸食K粉可导致男性吸食者出现性欲减退、阳痿、射精困难、早泄等性功能障碍;除此之外,精子数量减少或质量明显降低,进而对生育产生不利影响...

-
01/25
2021
细数冰毒的危害有哪些?
冰毒这种新型毒品对人危害极大:对人体中枢神经系统具有极强的刺激作用,且毒性强烈。冰毒具有极强的精神依赖性,吸食后会产生强烈的生理兴奋,大量消耗人的体力和降低免疫功...
